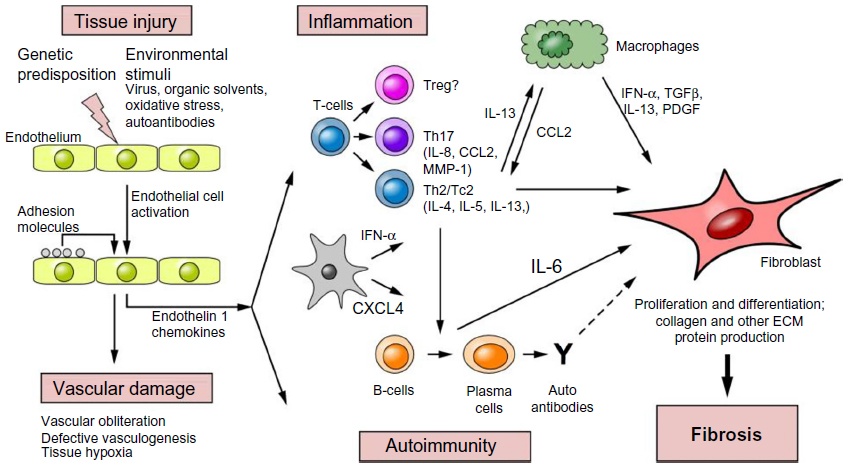
Topics: drug development, preclinical research, translational research, scleroderma, fibrosis, disease models

After decades of research into the genetics of cancer, there has been amassing evidence associating these cancer causing genes, or oncogenes, to cellular metabolism and bioenergetics regulation. This metabolic link serves to facilitate the rapid growth and survival that is characteristic of many cancer cells. Given this, it is not surprising that obesity, already known to exacerbate metabolic disorders, is also affiliated with increased cancer risk. As obesity approaches pandemic levels, understanding the mechanisms involved between cancer and organism metabolism becomes increasingly important.
Topics: cancer/tumor, AMPK
In its 2015 “Year-in-Review”, Science News declared the “Epigenome” has made its “debut”. Why? Because epigenetic alterations; including acetylation, phosphorylation and methylation have been found to be a means by which genes can be regulated post translation.
While not new, numerous clinical studies have recently been published highlighting the important role methylation plays in epigenetic alterations.
Published findings include:
Topics: Biomarkers, methylation, assays, epigenomic

Immunoinhibitory checkpoints — act as the BREAKS of an immune system — shutting down the adaptive immune response. While inhibitory checkpoints such as PD-1 and CTLA-4 are important in the long-term prevention of auto-immune disorders, cancer will often take advantage of these signaling molecules to avoid destruction by the host immune system. In light of this, development of antagonistic monoclonal antibodies to block these signaling molecules has led to the FDA approval of both an anti-CTLA-4 monoclonal antibody, ipilimumab, and two anti-PD-1 antibodies, pembrolizumab and nivolumab, for the treatment of cancer.
These immunotherapeutic treatments have demonstrated versatility in the treatment of multiple cancer types. Nivolumab — first approved for the treatment of metastatic melanoma in late 2014 — has since been approved for treating advanced lung cancer and renal cell carcinoma, further illustrating the versatility of this immunotherapeutic approach. While demonstrating promise as single agents, the efficacious effects of these therapies are dramatically improved when they are used in combination with other chemo-radiation, or immuno-therapies. This exciting area of therapeutic potential is still in its infancy and continues to grow as more and more is learned about harnessing the body’s own capabilities to destroy cancer.
Topics: cancer/tumor, Immuno-Oncology

St. Paul, Minn., June 24, 2016 - MD Biosciences launches the InnovX® research platform that’s designed to bridge the gap between preliminary, early stage and industry relevant life science product research discoveries by academic institute researchers, as well as de-risk necessary research investments.
Topics: Academic Research, Discovery, Discoveries, InnovX

The 2016 Arrowhead Translation Microbiome brought the who’s who of the microbiome community together last week in the heart of Boston. Top key opinion leaders in the field came together to discuss new therapeutic discoveries, disease indications and regulatory pathways. Seres Therapeutics, the leader in microbiome therapy development, opened the meeting discussing their pipeline and recently designated “FDA Breakthrough Therapy”, Ser-109, for the treatment of reoccurring C. difficile infections. Other big players in attendance were Commense and Osel, Inc, who are both working to understand the impact of the vaginal microbiome on health and disease and how to treat a dysbiotic vaginal environment.
Topics: Microbiome, translational

On Thusday, MD Biosciences' Lead Scientist Britnie James, Ph.D. presented at the 2nd Annual Translational Microbiome Conference held in Boston, MA. Her talk entitled "Understanding the Impact of Animal Vendors and the Microbiome in Tumor Progression and Treatment in a Model of Metastatic Breast Cancer" highlights the importance of the role of the micorbiome in disease progression and treatment.
Topics: Inflammation, cancer/tumor, Microbiome
As we wind down to years end, we at MD Biosciences would like to thank everyone, especially our collaborators, for making this year a success. We have undergone significant growth that we expect to continue throughout the upcoming year.
Topics: Pain, Inflammation, Neuro/CNS, post-operative pain, cancer/tumor, Microbiome, Biomarkers, CLIA, Neuropathy
As an already fully functioning good laboratory practices (GLP) operation, MD Biosciences has recently achieved a new stage of value through CLIA certification for our high quality-testing services in the clinical and diagnostic arenas. CLIA or the Clinical Laboratory Improvements Amendments was put in place at the federal level to ensure a high level of laboratory testing. Therefore, clients can be assured that MD Biosciences meets these high quality standards and feel confident that their samples will be handled with the utmost technical and professional care to obtain reliable results.
Topics: Personalized medicine, precision medicine, CLIA, Certification, High-quality

The 2015 annual AMP meeting opened Novemeber 5th in Austin TX, with a presentation by Dr Brian Druker titled “Imatinib as a Paradigm of Targeted Cancer Therapies.” This year’s recipient of the AMP Award for Excellence in Molecular Diagnostics, Dr Druker highlighted the importance of identifying patients through biomarkers and matching therapies to the patient. This theme continued through many of the presentations, posters and vendor exhibits the importance of providing physicians actionable results for patient care and therapeutics. Molecular diagnostics are critical to any discussion on precision or personalized medicine.
Topics: cancer/tumor, Microbiome, Biomarkers, Association for Molecular Pathology, Pathology, precision medicine





